
Polecamy Lista artykułów
|
Technika CO2Dwutlenek węgla w wodzie, najważniejsze reakcje, formy asymilacji węgla
Dwutlenek węgla spełnia bardzo ważną rolę – mianowicie dostarcza węgiel do budowy związków organicznych. Pobieranie go przez rośliny wodne związane jest z dużą różnorodnością form, w jakie przechodzi w środowisku wodnym. Formy występowania zależą od pH wody – może on przyjąć jedną z czterech form (wykres2, tabela 3). Rośliny potrafiące wykorzystywać CO2 związany w jonach wodorowęglanowych posiadają specyficzny enzym – anhydrazę węglanową umożliwiający przekształcanie jonów węglanowych w dwutlenek węgla (taki sam enzym zawierają w karboksysomach organizmy prokariotyczne – sposób ten wykorzystują do zwiększenia lokalnego stężenia CO2 w pobliżu miejsc karboksylacji rubisco) Asymilacja CO2 przez rośliny wodneDostępność CO2 dla roślin zanurzonych jest dość niska, swój udział w tym ma tutaj tempo dyfuzji gazów, prawie 10 tysięcy razy wolniejsze niż w powietrzu, przez co ogranicza tempo wzrostu roślin wodnych. Niewielka grubość liści typowa dla roślin zanurzonych znacznie zmniejsza znaczenie tego zjawiska. Również warstwa graniczna jest znacznie mniejsza – stanowi ją ok. 0,5 mm warstwa wody związana z powierzchnią liści stanowiącą barierę dyfuzyjną dla gazów. Kalkulator CO2/KH
Technika CO2 - najczęściej zadawane pytaniaTechnika CO2 - dwutlenek węglaPrzeczytaj nasze artykuły
Rodzaje trójników do rozdzielania CO2 
Bimbrownia - sposób dozowania CO2 dla początkującychBimbrownia CO2 opiera swoje działanie na produkcji dwutlenku węgla przez drożdże, które wytwarzają go w procesie fermentacji cukru.
3. Na przycięta plastikową końcówkę igły nakładamy wężyk akwarystyczny i wciskamy ją w otwór w nakrętce. 4. Następnie zalewamy miejsce połączenia silikonem akwarystycznym. 5. Nalewamy przygotowany wcześniej zacier do butelki za pomocą lejka. 6. Po nalaniu zacieru zakręcamy nakrętkę butelki jak najmocniej potrafimy. 8. Następnie z drugiej strony zaworka montujemy pozostałą część wężyka i doprowadzamy go do tzw. oczyszczalni. Jaka jest wydajność butli CO2? Co ile trzeba ją nabijać?Butla 1.5kg wystarcza: Musisz nabijać butlę częściej? Podejrzewasz nieszczelność swojej instalacji CO2?
Uwaga: są to autentyczne informacje potwierdzone od 2001r przez użytkowników naszego forum. Nie daj się nabrać sprzedawcom, którzy będą podawali Ci zawyżone dane!
Weź udział w dyskusjach na forum: Ile bąbelków na sekundę dozować w akwarium?Porady dotyczące nawożenia CO2 Właściwy poziom nawożenia CO2 zależy od warunków wzrostu roślin wodnych, liczby roślin oraz zapotrzebowania na CO2 poszczególnych roślin. Preparaty "CO2 w płynie" - czytaj więcejNa rynku pojawiła się ciekawa alternatywa dla drogich zestawów CO2 - nawozy zawierające "węgiel w płynie". Dyskutuj na forum o nawozie EasyCarbo - kliknij >> Rozpuszczanie dwutlenku węgla w wodzie
 Wbrew rozpowszechnionemu mniemaniu jest to sprawa bardzo prosta, gaz ten rozpuszcza się doskonale w wodzie ( czego doskonałym przykładem są napoje gazowane ! ), a panujące powszechnie przekonanie że tzw. reaktory to urządzenia skomplikowane ( i przez to drogie ) jest w zasadzie błędne. Sposób 1 - Dzwon W sprzedaży dostępne są małe dzwony firmy Tetra, w zestawie jest dzwon w kształcie walca wykonany z zielonkawego tworzywa, wąż, dwie przyssawki i butla z 11 g CO 2 . Dzwon wiesza się w akwarium i napełnia się dwutlenkiem węgla. Według producenta wystarcza to na potrzeby około 50 litrowego akwarium ( przy większym zbiorniku mozna dać więcej dzwonów ) jednak z mojego doświadczenia jest to dawka "fail safe" czyli nie zrobimy tym krzywdy, ale bardziej zarośnięte zbiorniki wymagają dużo więcej. Zalety : estetyka wykonania , Wady : dość wysoka cena jak za kawałek tworzywa. 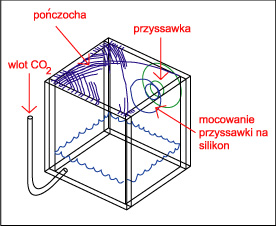 Sposób 2 - Dzwon własnej roboty Sposób 2 - Dzwon własnej roboty Do budowy dzwona będą nam potrzebne : szkło lub pleksi, klej do szkła lub pleksi, przyssawka z uchwytem ( np. od karmidełka lub zepsutego filtra ), pończocha o drobnych oczkach. Szkło lub pleksi sklejamy na kształt pudełka bez dna i przykrycia, na jednej ze ścian przyklejamy przyssawkę ( lub dwie, jeśli dyfuzor ma być większy ) a od góry smarujemy krawędź silikonem i wtapiamy pończochę, którą potem po wyschnięciu kleju przycinamy. Wymiary dzwona - wysokość około 5 cm ( większego parcia nie wytrzyma pończocha i gaz będzie uciekał ) a pozostałe wymiary należy dobrać doświadczalnie.Używam dzwona 5 na 3.5 na 3.5 cm w akwarium 4.5 l, używam też dzwona 13 na 5 na 5 cm w zbiorniku 100 l i takiego samego - z wymuszonym obiegiem wody na wylocie z kaskady - w zbiorniku 250 l. Skuteczność - do około 99 % Zalety : cena i możliwość dostosowania wielkości do własnych potrzeb, oraz to ze pończocha słuzy jako swego rodzaju zawór bezpieczeństwa - przy nadmiarze CO 2 parcie wody wypycha nadmiar gazu przez pończochę. Wady : zajmuje miejsce w akwarium i wymaga nakładu pracy na zbudowanie. Wadą wszystkich dzwonów jest to, że następuje dyfuzja gazów rozpuszczonych w wodzie do gazu zawartego w dzwonie - dzwon należy co pewnien czas opróżniać z gazu, a membranę oczyszczać z glonów. Sposób 2a - dzwon z wymuszonym obiegiem wody.  Konstrukcja dzwona jak wyżej , dodatkowo do układu podpinamy : Albo małą głowicę filtra, przy czym możemy zrobić podobnie jak proponuje Kolega tutaj, lub po prostu tylko skierować wylot z filtra na membranę ( mniejsza skuteczność ) Albo umieszczamy przy wylocie z kaskady. Możemy zbudować komin np. z pleksi lub polietylenu z butelki po napojach i przez taki komin skierować wodę PRZEZ membranę lub tak jak wyżej skierować strumień wody z filtra aby tylko omywał dzwon ( mniejsza skuteczność )  Albo montujemy na wylocie z filtra zewnętrznego. Całosc umieszczamy oczywiście pod wodą :) Zalety - jak w przypadku klasycznego dzwona, większa wydajność. Wady - jak w przypadku sposobu 2 Sposób 3 - podawanie CO2 do głowic filtrujących Elegancki i prosty sposób rozpuszczania nawet dużych ilości tego gazu. Podawać gaz można w kilka sposobów : Do rurki injektora jeśli nasz filtr taką posiada. Zalety - łatwe podłączenie. Wady - ponieważ CO 2 dość krótko przebywa w filtrze to nie rozpuszcza się w wodzie w 100 % - oczywiście nie zawsze jest potrzeba rozpuszczania dużych ilości gazu w wodzie ( np gdy produkcja bimbrowni przewyższa potrzeby ) . Drugą wadą jest to, że nie każdy lubi bąbelki w baniaku. Pod głowicę - należy jednak zaingerować w budowę filtra i albo wstawić między gąbkę a głowicę kawałek rurki z dziurką do której podłączamy rurke z CO 2 , próbować wcisnąć rurkę między gąbkę, głowicę i rurkę do mocowania gąbki lub po prostu dziurawiąc gąbkę. Obydwa sposoby raczej nie nadają sie do flitrów z obudową gąbki - należy ją albo zdjąć albo przedziurawić. Sposób skuteczniejszy niż powyższy, gdyż bąbelki są rozdrabniane przez wirnik. Zalety - wysoka skuteczność, Wady - gaz w trakcie rozpuszczania uderza o łopatki wirnika powodując hałas. 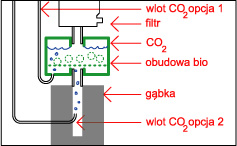 Do wkładu na filtr bio. Gaz podajemy jak w sposobie powyżej ( oczywiście przed wkładem a nie za ) . Dwutlenek węgla w zasadzie nie przeszkadza filtracji biologicznej, a że gaz przebywa dość długo w pudełku, to wzrasta skuteczność rozpuszczania. Do wkładu na filtr bio. Gaz podajemy jak w sposobie powyżej ( oczywiście przed wkładem a nie za ) . Dwutlenek węgla w zasadzie nie przeszkadza filtracji biologicznej, a że gaz przebywa dość długo w pudełku, to wzrasta skuteczność rozpuszczania. W przypadku, gdy nie potrzebujemy filtracji biologicznej ( a w holendrach często tak jest ) - po prostu wyjmujemy biosubstrat. Zalety - duża skuteczność i przy niezbyt dużym dawkowaniu CO 2 gaz ten nie uderza o wirnik. 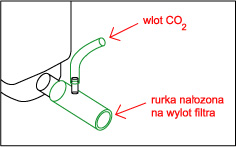 Do własnoręcznie wykonanej rurki injektorowej jeśli nasz filtr jej nie posiada, lub nie chcemy podawac CO 2 na wirnik. W rurce ( np ze starej deszczowni ) wykonujemy otwór do którego podłączamy gaz, całość mocujemy na wylocie filtra. Zalety - brak hałasu powodowanego przez bąbelki na wirniku, wady - rozpuszczanie nie jest tak skuteczne jak wtedy gdy gaz przechodzi przez wirnik. Uwaga ! ze względu na wysoką wydajność sposopów powyżej należy uważać przy podawaniu CO 2 z butli, przy przypadkowym odkręceniu zaworu można doprowadzić szybko do tragedii w zbiorniku !!! Sposób 4 - podawanie CO 2 do wlotu filtra kanistrowego. Również wydajny sposób nie zajmujący miejsca w akwarium; należy uważać na możliwość zapowietrzenia się filtra, a zakres stosowalności tej metody zależy od konkretnego modelu "kubełka" Sposób 4a - podawanie CO 2 do wlotu filtra kaskadowego. Sposób na rozpuszczenie stosunkowo małych ilości gazu z źródła ciągłego ( butla lub bimbrownia ), gdyż duże ilości gazu zapowietrzając rurkę unieruchomią filtr. Niektóre typy filtrów bardzo źle reagują na ten sposób. Sposób 5 - podawanie CO 2 na wylocie z filtra kanistrowego lub przelewowego otwartego. 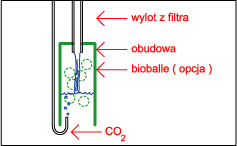 Należy skonstruować z plexi lub innego materiału coś w rodzaju kolumny, wypełnić ją np. bioballami, od dołu podawać dwutlenek węgla, a od góry wodę powrotną z filtra. Całość umieszczamy w akwarium. Sposób zaczerpnięty z thekrib.com ( moim zdaniem bioballe pomagają w rozpuszczaniu -zwiększają powierzchnię styku woda - CO 2 , ale nie są konieczne ! ) Rozwinięciem tej metody jest sposób 2a wypróbowany przez Autora. Należy skonstruować z plexi lub innego materiału coś w rodzaju kolumny, wypełnić ją np. bioballami, od dołu podawać dwutlenek węgla, a od góry wodę powrotną z filtra. Całość umieszczamy w akwarium. Sposób zaczerpnięty z thekrib.com ( moim zdaniem bioballe pomagają w rozpuszczaniu -zwiększają powierzchnię styku woda - CO 2 , ale nie są konieczne ! ) Rozwinięciem tej metody jest sposób 2a wypróbowany przez Autora. Sposób 6 - przez tzw "kamień" do napowietrzania Dość prymitywna metoda; jednak może być skuteczna gdy mamy do czynienia z małym akwarium i dużą produkcją CO 2 z bimbrowni. Warto stosować kamienie dające drobne bąbelki - np. takie z drewna stosowane w odpieniaczach. Zalety : cena i prostota, wady - mała skuteczność Sposób 7 - filtr plus jakiś pojemnik został omówiony przez Kolegę tutaj. Sposób 8 - rynna ze szkła Za czasopismem Akwarium, nr 1/90 s.15 ( artykuł pp K.Pająka i D.Pyrchli ) - szklaną rynnę o wymiarach około 5 na 5 cm, długości całego zbiornika, wklejamy kilka centymetrów nad dnem na tylnej ścianie. CO 2 dozujemy dwa razy dziennie. Rozwinęciem tej metody może być wykorzystanie podłużnego wzmocnienia akwarium i doklejenie do niego kilkucentymetrowego paska szkła, tak aby powstała rynna oddzielona od atmosfery. Sposoby dość kłopotliwe w montażu i zajmują dużo miejsca. Poza tym, tak jak dzwony , rynny wymagają okresowego oczyszczania z gazów dyfudujących z wody. Prosty kalkulatorek oznaczający zawartość CO2 w wodzie jest tutaj Jeśli Twoje rosliny wyglądają tak jak na obrazku na górze strony to wszystko z rozpuszczaniem CO2 jest w porządku ! Strona 4 z 7  |
| Działamy od 2001 roku i wspólnie z ponad 30 tysiącami akwarystów z całej Polski zdobywamy wiedzę i dzielimy się doświadczeniem oraz informujemy o nowościach z branży akwarystycznej. |






